
https://petroglifosrevistacritica.org.ve/
Identificación y caracterización de dos cepas causantes del hereque (Ralstonia solanacearum raza 2) en musáceas (AAA) del estado Aragua, Venezuela
Irmarú Z. Torres Perdomo1
Maximiliano Méndez Morales2
Glenda Y. Aponte Parra3
Mingrelia J. España Zarate1
José D. Rosales Rangel1
Sandy L. Molina Moret1
2Facultad de Agronomía, Universidad Central de Venezuela (UCV), Av. Universidad, vía El Limón. Maracay, Venezuela.
3Profesional de Investigación, Unidad de Protección Vegetal Instituto Nacional de Investigaciones Agrícolas, Centro Nacional de Investigaciones Agropecuarias (INIA - CENIAP). Av. Universidad, vía El Limón, Edif. INIA, Código Postal: 2105. Maracay, Venezuela.
La especie Ralstonia solanacearum (Smith) Yabuuchi, raza 2, causante del hereque o moko bacteriano en las musáceas, constituye una unidad taxonómicamente compleja que exhibe una amplia diversidad genética, fisiológica, serológica y de rango de hospedantes. Es por ello que se planteó identificar y caracterizar dos cepas (R4 y R5) seleccionadas por su patogenicidad. Para esto, se procesaron 27 muestras de 10 plantas sintomáticas de las que se obtuvieron colonias de forma irregular, elevación plana, aspecto mucoide, bordes gruesos, color crema y pigmentación central rojiza de diferentes intensidades, usando medios de cultivo selectivos como el agar nutritivo y tetrazolio. Las observaciones al microscopio determinaron la presencia de colonias puras, caracterizadas como Gram Negativas con bastones rectos y curvos. Las pruebas de patogenicidad realizadas en plantas sanas, inoculadas con las dos cepas por infiltración, dieron como resultado la sintomatología característica de la enfermedad. Para la identificación molecular se utilizaron los iniciadores universales U1 y U2, los cuales amplificaron una banda de 996 pb que, al secuenciarla y compararla con la base de datos del GeneBank®, revelaron 80,58% de identidad y 92% de cobertura en el aislado R4, y 81,74% de identidad y 82% de cobertura para el aislado R5. Además, se utilizaron los iniciadores específicos, OLI1 y Y2 que amplificaron una banda de 288 pb lo que permitió identificar las muestras estudiadas como R. solanacearum. Estos aislados podrán ser utilizados en distintos programas de mejoramiento para buscar tolerancia al mencionado patógeno y generar estrategias para el manejo integrado de la enfermedad.
The species Ralstonia solanacearum (Smith) Yabuuchi, race 2, which causes bacterial hereque or moko in musaceae, constitutes a taxonomically complex unit that exhibits a wide genetic, physiological, serological and host range diversity. For this reason, two strains (R4 and R5) selected for their pathogenicity were identified and characterized. For this, 27 samples from 10 symptomatic plants were processed, from which colonies of irregular shape, flat elevation, mucoid appearance, thick edges, cream color and reddish central pigmentation of different intensities were obtained, using selective culture media such as nutrient agar and tetrazolium. Microscopic observations determined the presence of pure colonies, characterized as Gram-negative with straight and curved rods. Pathogenicity tests performed on healthy plants inoculated with the two strains by infiltration resulted in the characteristic symptomatology of the disease. For molecular identification, the universal primers U1 and U2 were used, which amplified a 996 bp band that, when sequenced and compared with the GeneBank® database, revealed 80.58% identity and 92% coverage in isolate R4, and 81.74% identity and 82% coverage for isolate R5. In addition, the specific primers OLI1 and Y2 were used, which amplified a band of 288 bp, allowing the identification of the samples studied as R. solanacearum. These isolates can be used in different breeding programs to search for tolerance to this pathogen and to generate strategies for the integrated management of the disease.
Introducción
El hereque o moko bacteriano causado por Ralstonia solanacearum raza 2, es considerado uno de los principales problemas fitosanitarios que afectan a las musáceas en las regiones tropicales y subtropicales del mundo, y constituye un problema potencial para aquellos países o áreas en los que está presente debido a que afecta todos los estados de desarrollo de la planta, siendo un factor determinante en la restricción comercial de la producción (Belalcázar et al., 2004), ya que presenta un amplio rango de hospedantes, fácil diseminación, alta variabilidad genética y difícil manejo (Valencia et al., 2014). Por tanto, los problemas presentes en el manejo de la enfermedad y en los programas de mejoramiento para obtener cultivos resistentes, se debe a la amplia diversidad y a la distribución geográfica del agente etiológico; así como a la alta capacidad de adaptación a diferentes ambientes y hospedantes (Genin, 2010; Prieto Romo et al., 2012; Huet, 2014; Yuliar et al., 2015), por lo que si no se presenta un manejo adecuado del cultivo puede llegar a destruir en un 100% las plantaciones donde se presenta (Álvarez et al., 2015).
Esta especie bacteriana se clasifica en cinco razas, según la patogenicidad y el rango de plantas hospedantes que afecta (Prieto Romo et al., 2012). Asimismo, la clasificación en biovares se basa en la capacidad metabólica de utilizar tres disacáridos (celobiosa, lactosa y maltosa) y tres alcoholes hexosas (dulcitol, manitol y sorbitol) (Genin, 2010; Prieto Romo et al., 2012).
La raza 1 está principalmente relacionada con plantas pertenecientes a la familia Solanaceae, mientras que la raza 2 afecta a las familias Musaceae y Heliconiaceae en las que causa marchitez bacteriana en plátanos triploides y heliconias en áreas tropicales, denominándose Moko bacteriano. La raza 3, por su parte, afecta algunas solanáceas y geranios, principalmente en zonas templadas o de clima frío moderado tropical, la raza 4 afecta preferentemente al cultivo del jengibre (Genin, 2010; Prieto Romo et al., 2012; Peeters, 2013) y la raza 5 se especializa en plantas del género Morus sp. (European and Mediterranean Plant Protection Organization [EPPO], 2004).
Ha sido reportada como una de las más antiguas estudiada por los fitopatólogos (Rorer, 1911), y los avances científicos han logrado la secuenciación completa de su genoma (Salanoubat et al., 2002), demostrándose una gran variabilidad genética (Poussier et al., 2003; Yu et al., 2003) que le permite sobrevivir en diversas condiciones ambientales (Caruso et al., 2005). Es importante destacar, que para el aislamiento in vitro de la bacteria se debe recurrir a medios de tetrazolio (TZC) (Kelman, 1954) y las técnicas de metabolismo con disacáridos y alcoholes hexosa son una excelente estrategia para caracterizar al patógeno con relación al biovar (Hayward, 1964; French et al., 1995).
Las musáceas por su condición de ploidía son estériles, por lo que el mejoramiento genético tradicional tiene poca probabilidad de éxito. Dentro de las alternativas de mejoramiento para plantas de reproducción asexual, las mutaciones radio inducidas son una herramienta con posibilidades de éxito, sobre todo si se acopla a los sistemas de regeneración in vitro, técnicas que ya están a disposición de los investigadores y que en los últimos años han demostrado ser eficientes en el mejoramiento genético de los cultivos de interés agrícola (Salazar, 2014). De allí la importancia de tener el agente causal de la enfermedad bien caracterizado con el fin de iniciar la búsqueda de tolerancia a la enfermedad en genotipos radio inducidos.
La identificación de bacterias patógenas de plantas se puede realizar mediante el uso de métodos morfológicos, bioquímicos y moleculares (Schaad et al., 2001). Los dos primeros métodos permiten observar la morfología de la bacteria y de las colonias que forma, y las particularidades dentro de los distintos tipos bacterianos, que dan lugar a diferentes respuestas en los patrones de comportamiento metabólico, fisiológico y morfológico de los miembros de cada grupo (Apolinario Carmona, 2018).
Ahora bien, los métodos moleculares se han erigido como procedimientos confiables para la identificación de bacterias patógenas, dentro de los cuales el análisis de la secuencia génica del ARNr 16S es la herramienta más utilizada. Este marcador está presente en todas las bacterias como una familia de multigenes u operones cuya función no se modifica con el tiempo y actúa como un marcador eficiente de evolución (Fernández et al., 2010).
Las técnicas de diagnóstico han evolucionado de manera tal, que se han alcanzado niveles de detección rápidos y confiables permitiendo incluso, la detección asintomática en material de siembra sexual o asexual y de inóculo en sitios de siembra; puntos críticos para cualquier programa de manejo integrado de enfermedades (Kageyama et al., 2003). Tradicionalmente, la detección de R. solanacearum se ha basado en métodos como el aislamiento en medios de cultivo, pruebas de patogenicidad sobre plantas indicadoras y el uso de técnicas serológicas, especialmente del tipo ELISA (Robinson-Smith et al., 1995).
Cada uno de estos métodos presenta ventajas y desventajas relacionadas con sus niveles de sensibilidad y especificidad, rapidez y nivel de costos. Así, por ejemplo, las pruebas basadas exclusivamente en medios selectivos resultan económicas y técnicamente sencillas, aunque requieren de tiempos extensos y sus niveles de sensibilidad sólo alcanzan en promedio 104 UFC.g-1 de tejido o suelo. Por lo anteriormente expuesto, se hace necesario el uso de metodologías clásicas combinadas con métodos moleculares, que permitan una caracterización e identificación certera a corto plazo. Es por ello, que se planteó como objetivo de este trabajo identificar y caracterizar dos cepas bacterianas causantes del hereque en musáceas para ser utilizadas con fines de mejoramiento genético en la búsqueda de materiales tolerantes a R. solanacearum y también para evaluar estrategias en el manejo integrado de la enfermedad.
Materiales y Métodos
Muestreo
Se visitaron nueve fincas dedicadas al cultivo de musáceas ubicadas en el municipio Bolívar del estado Aragua, Venezuela. Se muestrearon 10 plantas sintomáticas de cambur (AAA) cultivar ´Pineo Gigante´, de las cuales se colectaron 27 muestras de diferentes órganos (hoja, pseudotallo, cormo). Las muestras se almacenaron por separado en nevera a 4°C hasta su análisis, en el Laboratorio de Bacteriología de la Unidad de Protección Vegetal del Instituto Nacional de Investigaciones Agrícolas (INIA).
Aislamiento y obtención de cultivos puros
Se pesó 1 g de tejido afectado para cada muestra, se esterilizó con alcohol isopropílico 70% por 2 min y se maceró en agua destilada con buffer Tris HCL pH 7,6. Para verificar la presencia de flujo bacteriano, se colocó una gota de este macerado en un portaobjeto y se observó en lupa estereoscópica de luz marca Leica modelo DM3000, utilizando el objetivo de 40X. Luego, en campana de flujo laminar con apoyo de un mechero, se realizó el sembrado en medio semi-selectivo, que contiene 1 g cloruro de TZC, 18 g Agar- Agar, 10 g dextrosa, 10 g peptona, 1 g casaminoacidos, 1 g de cristal violeta, diluidos en 1000 mL de agua destilada (Kelman, 1954; French et al., 1995; Denny y Hayward, 2001; Gómez et al., 2004). La siembra se realizó con un asa de platino, utilizando el método de estriado por agotamiento y se incubó por 72 h a ± 27°C. Pasadas las 48 h se marcaron las colonias, seleccionando las muestras que mostraron mayor intensidad en la coloración rojiza.
Caracterización morfológica y bioquímica
Las colonias purificadas fueron sometidas a diferentes pruebas para determinar sus características morfológicas y bioquímicas según la metodología planteada por Schaad et al. (2001). Para la caracterización morfológica se utilizó un microscopio de luz marca Leica modelo DM300 con un objetivo 100X, colocando una gota de aceite de inmersión. Se evaluaron las características de forma, elevación, bordes, color y aspecto de las colonias, y también se realizó la prueba de Tinción Gram. Para la caracterización bioquímica, se realizó la prueba de óxido-fermentación donde se determinó el requerimiento de oxígeno, para lo cual se utilizó el medio de Hugh y Leifson propuesto por Schaad et al. (2001), en el cual se inocularon bacterias con 24 h de crecimiento mediante el método picadura vertical profunda.
Postulados de Koch
Previo a la inoculacion de las plantas se preparó la solucion bacteriana, donde se procedió a cultivar la cepa en medio TZC (Kelman, 1954). Una vez crecida la colonia, se agregaron 5 mL de agua destilada estéril para lavar la superficie del medio, y a partir de allí se hicieron diluciones seriadas hasta obtener la concentracion adecuada a 1×10-8 UFC, siguiendo la escala propuesta por Mcfarlant (McFarlant, 1907; Klement et al., 1990) y corroborada con un bioespectrofotómetro Eppendorf Ag, modelo Hamburg 22331, a una densidad óptica de 0,1 y una longitud de onda de 600 nm (He et al., 1983; Hernández-Garboza y Bustamante-Rojas, 2001).
Se utilizaron cinco plantas provenientes del cultivo in vitro, las cuales fueron sometidas a una hora de luz UV previo a la inoculación. Una de estas plantas se utilizó como testigo infiltrando agua destilada estéril; dos plantas fueron infiltradas con un inóculo de la bacteria a 1×10-8 UFC, utilizando una mini inyectadora, una en la nervadura central de la segunda hoja más joven y la otra en la base del pseudotallo (Vivas et al., 2009). A las dos plantas restantes se les causó lesiones con un raspado suave en el envés de la segunda hoja utilizando carborundum® y se le aplicó el inóculo con un algodón impregnado del concentrado de bacterias. Las plantas se llevaron a una cámara húmeda y al observarse la aparición de los síntomas característicos del moko o hereque se procedió al reaislamiento siguiendo la metodología antes descrita.
Caracterización molecular
La extracción del ADN de las cepas se realizó según el protocolo de Laird et al. (1991). Se realizó la amplificación de la región ribosomal ADNr 16S con los iniciadores universales propuestos por Lu et al. (2000) para la identificación de procariontes: U1 (5′-CCA GCA GCC GCG GTA ATA CG-3′) y U2 (5’-ATC GG(C/T) TAC CTT GTT ACG ACT TC-3’). La reacción de amplificación se realizó en un volumen final de 40 µL conformada por tampón de PCR 1X; 2,5 mM de MgCl; 0,1 μM de cada iniciador, 1U de taq polimerasa, 0,2 mM de los dNTPs y 5 ng de ADN. La amplificación por PCR inició con una desnaturalización de 95ºC por 5 min, seguida de 40 ciclos de 95ºC por 30 s, 55ºC por 30 s y 72ºC por 30 s, culminando con una extensión final de 10 min a 72ºC.
La electroforesis se realizó en geles de agarosa al 1,5% con 5 mL del producto amplificado por pozo, y comparados con un marcador de peso molecular de 1 Kb plus Invitrogen®. Se utilizó una cámara horizontal marca BIORAD con buffer de corrida TBE 1X a 90 V durante 60 min. Una vez culminada la corrida electroforética, los geles fueron teñidos con SYBR® GREEN y visualizados con luz UV en el digitalizador de imágenes marca UVitec utilizando el software Alliance Vs. 15.05.
Posteriormente, se realizó una amplificación con iniciadores específicos para R solanacearum propuestos por Seal et al. (1993): OLI1 (5’-GGGGGTAGCTTGCTACCTGCC-3’) y Y2 (5’-CCCACTGCTGCCTCCCGTAGGAGT-3’). La reacción de amplificación se realizó en un volumen final de 10 µL, conformada por tampón de PCR 1X; 1,5 mM de MgCl; 0,5 μM de cada iniciador, 1U de taq polimerasa, 0,2 mM de los dNTPs y 10 ng de ADN. La amplificación por PCR inició con una desnaturalización de 96ºC por 5 min, seguida de 35 ciclos de 96ºC por 20 s, 67ºC por 25 s y 72º por 30 s, culminando con una extensión final de 10 min a 72ºC. La electroforesis y visualización de los productos de PCR se realizó de la misma manera que para la región ribosomal.
Análisis de las secuencias
Los productos resultantes de las amplificaciones fueron purificados y secuenciados. La secuenciación se realizó en la Unidad de Polimorfismo Genético de la Fundación Instituto de Estudios Avanzados (IDEA). Las secuencias se procesaron con el apoyo de herramientas de bioinformática, utilizando los programas Chromas Lite, versión 2.1.1 y BIOEdit 7.2.5, con el fin de generar las secuencias consenso y compararlas con las disponibles en el registro nucleotídico del Genbank (base de datos del National Center for BIOtechnology Information) (NCBI) utilizando la aplicación Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1990).
Resultados y Discusión
Caracterización morfológica y bioquímica
Esta caracterización permitió observar una morfología de colonias de aspecto mucoide, con forma irregular, elevación plana, con bordes gruesos color crema y pigmentación central rojiza de diferentes intensidades. Esta coloración rojiza permitió la selección de las muestras R4 y R5 por presentar la mayor intensidad, lo que sugiere mayor virulencia (Akter et al., 2021; Saquicela Cruz et al., 2023). Al observar estas cepas al microscopio de luz y utilizando un lente de inmersión, se apreció un color rojizo por la tinción de Gram, evidenciándose la presencia de bacterias Gram negativas (figura 1).
Asimismo, se observaron colonias puras, bastones rectos o curvos con dimensiones de 0,5 a 1,0 x 1,0 a 3 µm. Todas estas características coinciden con las reportadas por diversos autores, en sus trabajos de caracterización e identificación de R. solanacearum. (Agrios, 1988; Apolinario Carmona, 2018; Akter et al., 2021; Saquicela Cruz et al., 2023).
Figura 1. Coloración rojiza de las células de bacterias Gram negativas.

Nota. A: Se observan bordes gruesos color crema y pigmentación central rojiza. B: Colonias gran negativas con bastones rectos y curvos colores rojizos.
La prueba de Oxido-Fermentación con el medio Hugh y Leifson, confirmó la actividad aeróbica lo cual se ajusta a las características de R. solanacearum. En este caso particular es aeróbica, dada la coloración amarillenta observada en la superficie del tubo de la izquierda, mientras que en el tubo de la derecha, sellado para evitar el intercambio de oxígeno, no ocurrió reacción alguna (figura 2). Esta coloración coincide con lo reportado por MacFaddin (2003) y Saquicela Cruz et al. (2023), quienes explican que esto se debe a que algunas bacterias son capaces de metabolizar un hidrato de carbono por vías fermentativas, en este caso la glucosa, que es hidrolizada en dos moléculas de triosa, luego son convertidas a una cantidad de compuestos de 1 a 4 carbonos; donde el principal intermediario es el ácido pirúvico.
Figura 2. Prueba de óxido fermentación.

Nota. Obsérvese en el tubo de la izquierda, como el color amarillento se manifiesta en la parte superficial.
Postulados de Koch
Ambos métodos de inoculación, infiltración con una mini jeringa y heridas causadas con carborundum®, utilizados en este ensayo resultaron ser eficientes. La aparición de los síntomas característicos del moko o hereque, como la flacidez y el color amarillento en los bordes de las hojas desplazándose hacia la nervadura central, se observó a las 48 h en las plantas inoculadas por ambos métodos (figura 3), acentuándose en los días siguientes en todas las hojas que fueron inoculadas con la bacteria. Se logró el reaislamiento del agente causal y su identificación, coincidiendo con el aislado original.
Figura 3

A: Síntomas a las 48 horas de las plantas inoculadas por infiltración
B: Testigo sin inocular.
Se pudo observar que la inoculación por infiltración de la nervadura central con una mini jeringa, fue mucho más efectiva con relación a la aparición de los síntomas y dispersión en el resto de la planta (figura 4), esto se explica porque el modo de diseminación del patógeno es a través de los haces vasculares. La aparición temprana de los síntomas en plantas inoculadas, independientemente del método de inoculación, puede atribuirse a que el inóculo provenía de un primer aislamiento y purificación de las muestras colectadas de plantas sintomáticas, pues según Vivas et al. (2009), R. solanacearum cuando es mantenida y replicada varias veces en medios de cultivo con TZC atenúa su grado de patogenicidad, lo que hace que al ingresar en plantas sanas se comporte de manera “avirulenta”.
Gómez et al. (2004), reportaron poco éxito en la inoculación con R. solanacearum en cultivares de plátano al manifestarse los síntomas a partir del día 10 y también concluyen que el ambiente in vitro y la variabilidad genética influye en la bacteria tornándolas “avirulentas”. A esta misma conclusión han llegado otros autores como Robertson et al. (2004) y Castillo y Greenberg (2007). Es importante destacar, que esta prueba de inoculación y evaluación de los postulados de Koch, permitió comprobar la virulencia y patogenicidad de los dos aislados seleccionados e identificados como R4 y R5.
Figura 4. Plantas inoculadas por infiltración presentando síntomas característicos de la enfermedad a las 48 h.

Caracterización e identificación molecular
La amplificación de la región ribosomal ADNr 16S
La amplificación de los ADN de los dos aislados seleccionados (R4 y R5) con los iniciadores universales propuestos U1 y U2 generó una banda intensa de 996 pb (figura 5), tamaño esperado que coincide con el reportado por Alvez et al. (2011) y Apolinario Carmona (2018).
Figura 5. Amplificación del ADNr 16S (U1-U2) de la bacteria Ralstonia solanacearum en gel de agarosa al 1,5%.

Nota. M: Marcador 1Kb plus Invitrogene®, 1-3 Cepa R4. 4-6: Cepa R5. 7-8: Controles negativos. 9: Marcador de cuantificación Lambda (100 ng.µL-1).
Análisis de secuencias
Los datos obtenidos del BLAST, muestran un 80,58% de identidad con un 92% de cobertura en la muestra R4, mientras que en la R5 la identidad fue de 81,74% con un 82% de cobertura, correspondiéndose con R. solanacearum. Esta aproximación a la identificación puede explicarse dado el tamaño parcial de la secuencia obtenida, la cual fue de 996 pb.
La identificación molecular de especies bacterianas no es fácil, diversos autores mencionan que la identificación de especies; así como la correcta asignación taxonómica en bacterias se basa en los niveles de similitud de la secuencia completa del gen ADNr 16S (1550 pb). Sin embargo, aun cuando se logre amplificar completamente este gen pueden presentarse errores, produciendo secuencias de baja calidad que no permitan una completa identificación (Ebili et al., 2017; Troncoso, 2022).
Es por ello que para lograr una correcta identificación de la especie, muchos autores recomiendan la amplificación y secuenciación de múltiples regiones como los genes 23S ADNr, espaciador Inter génico transcripto (ITS), genes de poligalacturonasa (pgl), endoglucanasa (egl), hrp, hrpB, el gen de la proteína Kfra, entre otros (Obrador, 2016; Truke Arango, 2018; Troncoso, 2022).
Amplificación con iniciadores específicos para R. solanacearum
Como resultado de la amplificación en las dos muestras estudiadas, se obtuvo el fragmento esperado de 288 pb (figura 6), que corresponde a una región del gen ADNr 16S específica para R. solanacearum, lo que permitió identificar ambos aislados.
Figura 6. Amplificación del ARNr 16S con los cebadores OLI1 y Y2 en gel de agarosa al 1,5%.
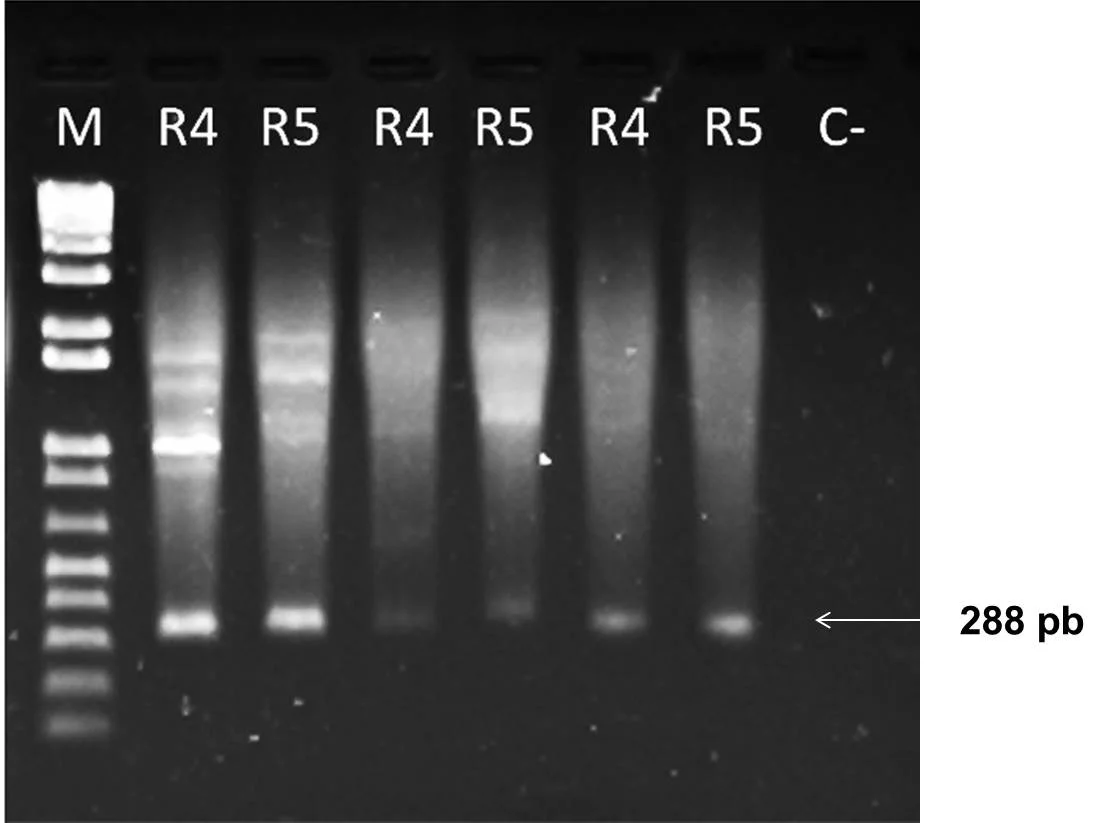
Nota. Se indica la banda característica de la bacteria Ralstonia solanacearum de 288 pb. M: Marcador 1Kb plus Invitrogene®. R4: Muestra cepa R4. R5: Muestra cepa R5. C-: Control negativo.
Conclusión
Una vez realizadas todas las pruebas morfológicas, bioquímicas y moleculares conducentes a la correcta identificación del patógeno causante de la enfermedad moko o hereque bacteriano en las musáceas (AAA) Cv ´pineo gigante´, se concluye que los aislados R4 y R5 corresponden con la bacteria Ralstonia solanacearum. Estos aislados serán incorporados al programa de mejoramiento genético de la Fundación IDEA, para buscar tolerancia al mencionado patógeno y generar estrategias para el manejo integrado de la enfermedad en el cultivo de las musáceas.
Agrios, G. N. (1988). Plant Pathology (3°ed). Academic Press.
Akter, N., Islam, M., Hossain, M., Islam, M., Chowdhury, S., Hoque, S., Nitol, R., y Tasnin, R. (2021). Management of wilt complex of eggplant (Solanum melongena L.) caused by Fusarium oxysporum, Ralstonia solanacearum and Meloidogyne spp. American Journal of Plant Sciences, 12 (1), 1155-1171. https://doi.org/10.4236/ajps.2021.127080
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., y Lipman, D. J. (1990). Basic local alignment search tool. Journal of Molecular Biology, 152 (3), 403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Álvarez, E., Pantoja, A., Gañán, L., y Ceballos, G. (2015). Current status of Moko disease and the Caribbean, and options for managing them. Centro Internacional de Agricultura Tropical (CIAT), Food and Agriculture Organization of the United Nations (FAO). https://www.fao.org/publications/card/en/c/d120154e-f795-432d-853a-64a4a2338fa7/
Alvez, B., Carballo, J., Alonso, G., y Oropeza, M. (2011). Diagnóstico molecular de Xanthomonas albilineans (Ashby) Dowson en Venezuela, agente causal de la escaldadura foliar de la caña de azúcar. Agronomía Tropical, 61(1), 27-36. http://ve.scielo.org/scielo.php?script=sci_arttext&pid=S0002-192X2011000100003
Apolinario Carmona, G. G. (2018). Identificación bioquímica y molecular de bacterias patógenas aisladas de plantas de papa (Solanum tuberosum) cultivadas en Chirgua, Edo. Carabobo, Venezuela [Tesis de grado, Universidad Central de Venezuela]. http://saber.ucv.ve/bitstream/10872/19685/1/TEG%20Genesis%20Apolinario.pdf
Belalcázar, C. S. L., Rosales, F. E., y Pocasangre, L. E. (2004). El “Moko” del plátano y banano y el rol de las plantas hospederas en su epidemiología. Memorias. XVI Reunión Internacional Acorbat (2004), Publicación especial, 16-35. http://publico.senasica.gob.mx/includes/asp/download.asp?IdDocumento=23605&IdUrl=78503&objeto=Documento&IdObjetoBase=23605&down=true
Castillo, J. A., y Grennberg, J. T. (2007). Evolutionary dynamics of Ralstonia solanacearum. Applied and Environmental Microbiology, 73 (4), 1225-1238. https://doi.org/10.1128%2FAEM.01253-06
Caruso, P.; Palomo, J. L., Bertolini, E., Álvarez, B., López, M. M., y Bioscan, E. G. (2005). Seasonal variation of Ralstonia solanacearum Biovar 2 populations in a Spanish river: Recovery of stressed cells at low temperatures. Applied and Environmental Microbiology, 71 (1), 140-148. https://doi.org/10.1128%2FAEM.71.1.140-148.2005
Denny, T., y Hayward, A. (2001). Gram – negative bacteria. En N. W. Schaad, J. B. Jones y W. Chum (eds). Laboratory guide for identification of plant pathogenic bacteria (pp. 151-174). American Phytopathological Society. https://www.scirp.org/(S(351jmbntvnsjt1aadkozje))/reference/ReferencesPapers.aspx?ReferenceID=71425
Ebili, H. O., Hassall, J. C., Fadhil, W., Ham-Karim, H., Asiri, A., Raposo, T. P., Agboola, A. J., y Ilyas, M. (2017). “Squirrel” Primer-based PCR assay for direct and targeted sanger sequencing of short genomic segments. Journal of Biomolecular Techniques, 28(3), 97-110. https://doi.org/10.7171%2Fjbt.17-2803-001
European and Mediterranean Plant Protection Organization. (2004). Ralstonia solanacearum. OEPP/EPPO Bulletin, 34,173-178. https://doi.org/10.1111/j.1365-2338.2004.00715.x
Fernández, A., García de la Fuente, C., Sáez, J., y Valdezate, S. (2010). Métodos de identificación bacteriana en el laboratorio de microbiología. Procedimientos en microbiología clínica. 37, 9-14. https://www.sciencedirect.com/science/article/pii/S0213005X11001571
French, E., Gutarra, L., Alley, P., y Elphinstone, J. (1995). Culture media for Ralstonia solanacearum isolation, identification and maintenance. Fitopatología Peruana, 30 (3), 126-130.
Genin, S. (2010). Molecular traits controlling host range and adaptation to plants in Ralstonia solanacearum. New Phytologist., 187, 920-928. https://doi.org/10.1111/j.1469-8137.2010.03397.x
Gómez, E. A., Álvarez, E., y Llano, G. (2004). Identificación y caracterización de cepas de Ralstonia solanacearum, raza 2, agente causal del moko de plátano en Colombia. Fitopatología Colombiana, 28 (2), 71-75. https://cgspace.cgiar.org/handle/10568/44341
Hayward, A. C. (1964). Characteristics of Pseudomonas solanacearum, Journal Applied Bacteriology, 27 (2), 265-277. https://doi.org/10.1111/j.1365-2672.1964.tb04912.x
He, L. Y., Sequeira, L., y Kelman, A. (1983), Characteristics of strains of pseudomonas solanacearum from China. Plant Disease, 67 (12), 1357-1361. https://worldveg.tind.io/record/600/
Hernández-Garboza, L., y Bustamante-Rojas, E. (2001). Control biológico de la marchitez bacterial en tomate con el uso de enmiendas orgánicas. Manejo Integrado de Plagas, 62,18-28. https://repositorio.catie.ac.cr/handle/11554/5964
Huet, G. (2014). Breeding for resistances to Ralstonia solanacearum. Front Plant Science, 5 (715), 1-5. https://www.frontiersin.org/articles/10.3389/fpls.2014.00715/full
Kageyama, K., Komatsu, T., y Suga, H. (2003). Refined PCR protocol for the detection of plant pathogens in soil. Journal of General Plant Pathology, 69,153-160. https://link.springer.com/article/10.1007/s10327-002-0037-4
Kelman, A. (1954). The relationship of pathogenicity in Ralstonia solanacearum to colony appearance on a tetrazolium medium. Phytopathology, (44), 693-695. https://www.cabdirect.org/cabdirect/abstract/19551101405
Klement, Z., Mevrides, A., Rudolph, K., Vidaver, A., Perombelon, M., y Moore, L. W. (1990). Inoculation of plant tissue. In methods in Phytobacteriology. P. 95-124. https://cir.nii.ac.jp/crid/1130282269926984192
Laird, P. W., Zijderveldl, A., Linders, K., Rudnicki, M. A., Jaenisch, R., y Berns, A. (1991). Simplified mammalian DNA isolation procedure. Nucleic Acids Research, 19 (15), 4293. https://doi.org/10.1093/nar/19.15.4293
Lu, J., Perng, C., Lee, S., y Wan, C. (2000). Use of PCR with universal primers and restriction endonuclease digestions for detection and identification of common bacterial pathogens in cerebrospinal fluid. Journal of Clinical Microbiology, 38, 2076–2080. https://doi.org/10.1128/jcm.38.6.2076-2080.20
MacFaddin, J. F. (2003). Pruebas bioquímicas para la identificación de bacterias de importancia clínica. Médica Panamericana. https://books.google.com/books?hl=es&lr=&id=FYWSzy7EjR0C&oi=fnd&pg=PA1&dq=MacFaddin,+J.+F.+(2003).+Pruebas+bioqu%C3%ADmicas+para+la+identificaci%C3%B3n+de+bacterias+de+importancia+cl%C3%ADnica.+Ed.+M%C3%A9dica+Panamericana.+850+p.&ots=RPSMRgP6Tt&sig=46pf2813M_R6qAStmYnhTzVaS7Y
McFarlant, J. (1907). The nephelometer: An instrument for estimating the members of bacteria in suspension used for calculating the opsonic index and for vaccines. Journal Applied Medical Association, 14, 1176-1178. http://dx.doi.org/10.1001/jama.1907.25320140022001f
Obrador, S. J. (2016). Variabilidad genética de la bacteria Ralstonia solanacearum de cepas aisladas de plátano en México. [Tesis Doctoral, Centro de Investigación Científica de Yucatán]. https://cicy.repositorioinstitucional.mx/jspui/bitstream/1003/468/1/PCB_BT_D_Tesis_2016_Obrador_Jose.pdf
Poussier, S., Thoquet, P., Trigalet-Demery, M., Barthet, S., Meyer, D., Arlat, M., y Trigalet, A. (2003). Host plant dependent phenotypic reversion of Ralstonia solanacearum from non-pathogenic to pathogenic forms via alterations in the phcAgene. Molecular Microbiology, 49 (4), 991-1000. https://doi.org/10.1046/j.1365-2958.2003.03605.x
Peeters N., Guidot A., Vailleau F., y Valls, M. (2013). Ralstonia solanacearum, a widespread bacterial plant pathogen in the post-genomic era. Molecular Plant Pathology, 14 (7), 651-662. https://doi.org/10.1111/mpp.12038
Prieto Romo, J., Morales Osorio, J. G., y Salazar Yepes, M. (2012). Identification of new hosts for Ralstonia solanacearum (Smith) race 2 from Colombia. Revista Protección Vegetal, 27 (3), 151-161. http://scielo.sld.cu/scielo.php?script=sci_arttex&pid=s1010- 27522012000300003
Robertson, A. E., Wetchter, W. P., Denny, T. P., Forthum, B. A., y Kluepfel, D. A. (2004). Relahionship between avirulence gene (avrA) Diversity en Ralstonia solanacearum and bacteria wilt incidence. Molecular Plant-Microbe Interactions, 17 (12), 1376-1384. https://doi.org/10.1094/MPMI.2004.17.12.1376
Robinson-Smith, A., Jones, P., Elphinstone, J. G., y Forde, S. M. D. (1995). Production of antibodies to Pseudomonas solanacearum, the causative agent of bacterial wilt. Food and Agricultural Immunology, 7 (1), 67-79. https://doi.org/10.1080/09540109509354866
Rorer, J. B. (1911). A bacterial disease of banana and plantains. Phytopathology, 1 (2), 45-49.
Salanoubat, M., Genin, S., Artiguenave, F., Gouzy, J., Mangenot, S., Arlat, M., Billault, A., Brottier, P., Camus, J. C., Cattolico, L., Chandler, M., Choisne, N., Claudel Renard, C., Cunnac, S., Demange, N., Gaspin, C., Lavie, M., Moisan, A., Robert, C., Saurin, W., Schiex, T., Siguier, P., Thébault, P., Whalen, M., Wincker, P., Levy, M., y Weissenbach, J., Boucher, C. A. (2002). Genome sequence of the plant pathogen Ralstonia solanacearum. Nature, 415, 497-502. https://www.nature.com/articles/415497a
Salazar, E. (2014). Radiaciones ionizantes en Musa AAA para inducción de mutaciones hacia la tolerancia a estrés hídrico [Tesis Doctoral, Escuela Socialista de Agricultura Tropical (ESAT)].
Saquicela Cruz, P. S., Romanova, E. V., Guamán Guamán, R. N., Ulloa Cortázar, S. M., y Villavicencio Abril, A. F. (2023). Caracterización morfológica y bioquímica de Ralstonia solanacearum Raza 2, bacteria patógena en cultivos de banano y plátano en El Carmen, Manabí, Ecuador. Siembra, 10 (1), e4305. https://doi.org/10.29166/siembra.v10i1.4305
Seal, S. E., Jackson, L. A., Young, J. P. W., y Daniels, M. J. (1993). Differentiation of Pseudomonas solanacearum, Pseudomonas syzygii, Pseudomonas pickettii and the blood disease bacterium by partial 16S rRNA sequencing: construction of oligonucleotide primers for sensitive detection by polymerase chain reaction. Microbiology, 139 (7), 1587-1594. https://www.microbiologyresearch.org/content/journal/micro/10.1099/00221287-139-7-1587
Schaad, N. W., Jones, J. B., y Chun, W., (2001). Laboratory guide for identification of Plant Pathogenic Bacteria (3°ed). Library of congress catalog card number 00-109131. https://www.cabdirect.org/cabdirect/abstract/20013064240
Troncoso, V. (2022). Propuesta metodológica para la caracterización molecular específica de Ralstonia solanacearum basada en el marcador molecular ADN ribosomal 16S [Tesis de Maestría, Universidad de Concepción]. http://repositorio.udec.cl/bitstream
Truke Arango, M. (2018). Caracterización patogénica y molecular de Ralstonia solanacearum raza 2, agente causal del Moko en plantaciones de plátano y banano (Musa spp.) en el Norte del Valle del Cauca [Tesis de Maestría, Universidad Nacional de Colombia]. https://repositorio.unal.edu.co/handle/unal/68913
Valencia, V. L., Álvarez, C. E., y Castaño, Z. J. (2014). Resistencia de treinta y cuatro genotipos de Plátano (Musa aab) y banano (Musa aaa) a cinco cepas de Ralstonia solanacearum raza 2 (Smith). Agronomía, 22 (2), 21 – 34. http://200.21.104.25/agronomia/download s/Agronomia 22(2)3.pdf.
Vivas, Y., Urdaneta, I., Rangel, S., y Hernández, J. (2009). Caracterización e incidencia de Ralstonia solanacearum Smith en plantas de Musa AAB en el Sector “El Roble”, Sur del Lago de Maracaibo, Venezuela. Revista UDO Agrícola, 9 (2), 383-392. http://udoagricola.orgfree.com/V9N2UDOAg/V9N2Vivas383.htm
Yu, Q., Alvarez, A. M., Moore, P. H., Zee, F., Kim, M. S., De Silva, A., Hepperly, P. R., y Ming, R. (2003). Molecular diversity of Ralstonia solanacearum isolated from gingering Hawai. Bacteriology, 93 (9), 1124 – 130. https://doi.org/10.1094/PHYTO.2003.93.9.1124
Yuliar, Nion, Y. A., y Toyota, K. (2015). Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbiology Environment, 30 (1), 1-11. https://doi.org/10.1264%2Fjsme2.ME14144